| О себе | Моя работа | Лечение | Библиотека | Дневник | Контакты | |||||||||
 | Аномальное сексуальное поведение(Под редакцией А. А. Ткаченко. М., 1997)Глава 7Состояние моноаминовых систем при парафилияхБ.М.Коган, А.З.Дроздов, А.А.Ткаченко, А.В.Талицкий, И.А.Ковалева, Т.С.Филатова, И.В.Маньковская, Л.О.Пережогин, Ю.Е.Куниковский7.1 Участие медиаторов в деятельности ЦНСРоль и значение катехоламинов в жизнедеятельности организма достаточно хорошо известны (Глебов Р.Н., Крыжановский Г.Н., 1978; Машковский В.Д. с соавт., 1983; Kopin, 1985; Kagedal B., Goldstein D.S., 1988; Nyyssonen, Parviainen, 1989; Esler et al., 1990;). Основными катехоламинами (КА) являются норадреналин (НА), адреналин (А), дофамин (ДА). Катехоламины осуществляют синаптическую передачу нервного импульса в центральной и вегетативной нервных системах, или, иными словами, являются нейромедиаторами. Помимо этого КА, в первую очередь адреналин, выполняют гормональные функции широкого спектра действия. Ядра НА-ергических систем головного мозга располагаются в продолговатом и среднем мозге (в частности, в синем ядре), а терминали оканчиваются практически во всех отделах головного мозга, включая подкорковые ядра, гипоталамус, мозжечок и кору головного мозга (Topel, 1985; Rogeness et al., 1992). Эти нейронные структуры модулируют и контролируют интегративную деятельность головного мозга, организуют согласованную активность практически всех компонентов центральных нервных механизмов. Дофаминовые системы головного мозга организованы в более компактные системы, выполняющие четко очерченные функции. Наиболее мощными ДА-ергическими системами являются нигростриальная, регулирующая моторные функции организма, мезолимбическая и мезокортикальная, играющие заметную роль в формировании мотивационных механизмов влечений; гипоталамические системы, контролирующие секрецию пролактина гипофизом и некоторые другие эндокринные функции (Topel, 1985; Rogeness et al., 1992). Долгое время считалось, что адреналин не является центральным нейромедиатором, однако к настоящему времени имеются предположения о существовании в мозге также и адреналинергических нейронов (Hokfelt et al, 1974). На периферии организма НА выступает в роли медиатора симпатической нервной системы, которая осуществляет контроль над всеми внутренними органами организма, включая кардиоваскулярную систему, желудочно-кишечный тракт, системы выделения и детоксикации. Адреналин является основным гормоном мозгового слоя надпочечников, который вместе с симпатическими структурами составляет симпатоадреналовую систему организма, являющуюся базой приспособительных адаптивных реакций организма, и обеспечивающую соответствующие гомеостатические перестройки в ответ на любые воздействия. Имеются косвенные данные, свидетельствующие, что на периферии организма, в частности в почках, могут существовать и дофаминергические нейроны, таким образом, ДА также выполняет нейромедиаторные функции и в периферической нервной системе (Ball et al., 1982; Bradley, Hjemdahl, 1986). Широта биологических функций катехоламиновых структур наряду с неспецифичностью и уникальной ролью этих молекул в гомеостатическом балансе организма чрезвычайно затрудняют исследование роли нарушений ключевых этапов метаболизма КА в патогенезе психических расстройств. Действительно, оценка особенностей центральных моноаминовых систем по результатам исследований периферических жидкостей является весьма непростой задачей. В качестве иллюстрации можно привести следующие примеры. Катехоламиновые и индоламиновые системы реагируют на все виды стрессов, включая физическую нагрузку и действие психологических раздражителей, системы адаптации меняют свой статус при практически любых соматических заболеваниях (Большакова Т.Д., 1973; Васильев В.Н., Чугунов В.С., 1985). Неврологические расстройства также в значительной мере связаны с деятельностью моноаминовых механизмов (Чугунов В.С., Васильев В.Н.,1984). Не вызывает никаких сомнений тот факт, что патология моноаминовых систем является одним из этиологических факторов многих психических нарушений. 7.1.2. Основные пути метаболизма медиаторовПринципиальные пути метаболизма катехоламинов в настоящее время достаточно хорошо изучены и информация об этом представлена в исчерпывающих обзорах, вышедших в последнее десятилетие. Системы утилизации активных нейромедиаторных и гормональных молекул имеют огромное значение для нормальной жизнедеятельности организма. Быстрое устранение действующих медиаторных единиц не менее важно, чем механизмы оперативного выброса нейромедиатров в ответ на нейрональный потенциал действия. Хорошо известно, что основными процессами инактивирования катехоламиновых медиаторов в синаптической щели являются энергозависимый специфический обратный захват НА и ДА пресинаптическим окончанием и метилирование медиаторных молекул, катализируемое катехол-О-метилтрансферазой. Моноаминоксидаза является важнейшим ферментом катаболизма КА. Считается, что имеются две основные формы моноаминоксидазы (МАО) — МАО-А и МАО-Б, различающиеся в первую очередь по субстратной и ингибиторной специфичности. Ферментная система не обладает узкой субстратной специфичностью и способна окислять широкий круг моноаминов и их метаболитов. Образовавшаяся в ходе реакции молекула альдегида весьма неустойчива и сразу подвергается окислению до кислоты или восстановлению до спирта. (Горкин В.З.,1981). По мнению большинства исследователей, основными ферментативными системами катаболизма катехоламиновых нейромедиаторов в организме являются моноаминоксидаза и катехол-О-метилтрансфераза (КОМТ). Однако известно также о существовании третьего пути биохимической инактивации катехоламинов — конъюгационных процессах с образованием их сульфо- и глюкуроновых эфиров. Изучению особенностей этого пути метаболизма катехоламинов уделялось значительно меньшее внимание, несмотря на то, что многие факты о биохимической стороне процессов образования и выведения конъюгатов относительно давно известны. Итак, в организме присутствует большой пул конъюгированных молекул как самих катехоламинов, так и их метаболитов. Феномен конъюгационного инактивирования КА широко распространен среди млекопитающих, и человек обладает одной из самых развитых систем конъюгирования КА, выполняющей важные физиологические функции (Dousa, Tyce, 1988). У людей около 70 % А и НА экскретируются в виде конъюгатов, в основном сульфатов (Kahane et al., 1967), в крови человека 95 — 99 % дофамина находится в форме сульфатов, концентрация конъюгированных форм НА, А, норметанефрина в 3-4 в раза превосходит показатели свободных форм этих молекул в циркуляции (Johnson et al., 1980; Yoneda et al., 1984). Конъюгированные формы катехоламинов (КА-К) составляют основную часть экскретируемых молекул КА (60 — 80 %), в первую очередь это относится к ДА (Van Loon, 1980; Scott, Elchisak, 1983, Yamamoto et al., 1995). Основные метаболиты КА — диоксифенилуксусная кислота (ДОФУК), гомованилиновая кислота (ГВК), ванилилминдальная кислота (ВМК), метанефрин (М), норметанефрин (НМ), метоксифенилэтиленгликоль (МОФЭГ) также экскретируются главным образом в виде конъюгированных форм (Roth J.A., Rivett J. 1982). Более того, коньюгаты КА, МОФЭГ и других метаболитов составляют большую часть этих веществ в спинномозговой жидкости. В то же время молекулы-предшественники катехоламинов в цепи биосинтеза активных медиаторов (диоксифенилаланин (ДОФА), тирозин) в меньшей степени подвержены реакциям конъюгирования (Roth J.A., Rivett J., 1982). Таким образом, конъюгационные процессы представляют универсальный механизм превращения молекул, имеющих катехольное кольцо, с образованием метаболических неактивных молекул, которые являются основной формой существования катехоламинов в биологических жидкостях человека. Кроме того, популяция НА-К более стабильна по сравнению со свободными формами и не столь резко меняется на фоне кратковременных стрессорных воздействий (Claustre et al., 1983; Ratge et al., 1986), тогда как более длительные периоды увеличения активности симпатической нервной системы могут приводить к увеличению концентрации связанных форм КА в крови (Gaudin et al., 1990). При циррозе, например, на фоне хронического подъёма уровня НА и ДА в плазме крови возрастает содержание и сульфатированной формы дофамина (ДА-С) и глюкуронированной формы норадреналина (НА-Г) (Gaudin et al., 1990). Однако существует и такое мнение, что активность симпатических нейронов не связана с уровнем КА-К в крови. Так, Cuche et al. (1990) продемонстрировали, что после сильных стрессорных воздействий (электрошок и инсулиновая гипогликемия) содержание КА-C в крови через 2 и 20 мин не увеличивается, а снижается, тогда как уровень НА-Г через 2 мин возрастает. Тем не менее, в связи с тем, что глюкурониды составляют небольшую часть КА-К, это не может играть существенной роли в метаболизме активных молекул медиаторов. Не вызывает сомнений, что уровень КА-К в крови является более стабильной популяцией молекул по сравнению с их свободными формами и уровень КА-К не подвержен быстрым изменениям при перестройках функционального состояния симпатической нервной системы. 7.1.3. Роль моноаминов в патогенезе психических расстройствПри изучении участия катехоламиновой системы в патогенезе психических и неврологических заболеваний возникает закономерный вопрос о роли инактивационного конъюгирования в синаптических процессах головного мозга. Суммируя известные литературные данные, можно утверждать, что популяция конъюгированных молекул катехоламинов отличается следующими чрезвычайно важными для поддержания гомеостаза особенностями. С одной стороны, сульфо- и глюкуроноконъюгирование является эффективным и быстрым механизмом для инактивации медиаторных и гормональных веществ как в мозге, так и на периферии. С другой стороны, связанные КА не являются биологически нейтральным пулом молекул и формируют депо биологически активных молекул. В заключение нужно отметить, что в настоящее время исследователи уже стали осознавать, что роль конъюгирования КА в организме (учитывая функции самих КА в поддержании гомеостаза) достаточно велика, хотя до сих пор прикладные работы в основном посвящены исследованию суммарного количества КА и их метаболитов. Несмотря на то, что в отдельных работах получены интересные результаты по содержанию КА-К при депрессиях (Jimerson et al., 1981; Kienzl et al., 1990; Mine et al., 1993), болезни Паркинсона (Kienzl et al., 1990), наркоманиях (Faraj et al., 1993) и других расстройствах, период активного прикладного использования накопленных теоретических данных о механизме конъюгирования КА при изучении биологических основ психических заболеваний еще не наступил. Состояние проблемы соотношения клинических признаков психических расстройств и особенностей обмена катехоламиновых медиаторов в настоящее время трудно назвать удовлетворительным. Сложилась весьма интересная ситуация — за 35 лет интенсивных исследований накоплен огромный фактический материал по данному вопросу, однако вследствие множества противоречивых данных единой концепции участия КА — систем в патогенезе аффективных расстройств до сих пор не создано, не выявлены прогностические и диагностические особенности метаболизма, позволяющие не только предсказывать риск возникновения или прогредиентность имеющихся расстройств, но и подбирать адекватные терапевтические средства. Парадоксальность ситуации усугубляется тем, что в начале систематических исследований моноаминовых систем при депрессиях вопрос казался более ясным, чем сейчас, поскольку клинические наблюдения свидетельствовали, что вещества, вызывающие истощение содержания КА в пресинаптических окончаниях (например резерпин), стимулируют проявление депрессии у людей. Кроме того, исследования in vitro продемонстрировали, что некоторые эффективные антидепрессанты являются блокаторами обратного захвата моноаминов. В основном на этих фактах базировались созданные концепции участия моноаминовых систем в патогенезе аффективных расстройств, появившиеся в 60-х годах. Так, в работах (Shildkraut 1965,1973., Bunney, Davis 1965) выдвигались гипотезы о недостаточности количества молекул НА в центральных синаптических контактах, тогда как другие авторы подчеркивали дефицит индоламиновой нейропередачи при депрессиях (van Praag H.M., Korf, 1971; Coppen, 1972). Однако, интенсивные биохимические исследования не смогли подтвердить или опровергнуть гипотезу о недостаточности моноаминовой нейромедиации как основной причины депрессивных проявлений. Сформулированная концепция о биохимической гетерогенности депрессий с существованием форм при которых преимущественно поражаются индоламиновые или катехоламиновые системы нейромедиации (van Praag H.M., Korf, 1971) также не привела к плодотворным результатам, поскольку последующие исследования опровергли предположения о существовании столь «чистых» форм депрессивных расстройств (van Praag H.M., Lemus, 1986; van Praag H.M. et al., 1990). По мнению, Rogeness et al.(1990), функционирование НА — систем головного мозга и периферической нервной системой снижено при расстройствах поведения асоциальной направленности и увеличено при депрессиях, тревожности, агарофобии и др. заболеваниях. Люди из первой группы, которые отличались наличием асоциального поведения, менее чувствительны к опасности, риску, общественному порицанию, у них отсутствуют проявления тревожности, они склонны к социально нежелательному поведению и преодолению социально установленных правил поведения. Люди, с выраженными депрессивными проявлениями, из второй группы, напротив, чрезмерно чувствительны к внешним воздействиям, к наказанию, социальному порицанию, чересчур возбудимы и тормозимы вследствие гиперфункций НА — систем. Исходя из этой теоретической предпосылки, авторы (Rogeness et al. 1990), сумели показать, что в группе детей с низкой активностью дофамин-бета-гидроксилаза (ДБГ) выше процент расстройств поведения, а в группе с высокой — больше тревожных расстройств и депрессивных проявлений. Активность ДБГ они рассматривали главным образом как индикатор интенсивности выброса медиатора в синаптическую щель и, таким образом, уровня функционального состояния симпатической нервной системы, хотя известно что нет прямой взаимосвязи между активностью ДБГ плазмы крови и состоянием симпатических нервов (Meltzer et al., 1976; Kopin et al., 1976; Sellers et al., 1978). Кроме того, соотношение МОФЭГ/ВМК и МОФЭГ/НА+НМ+ВМК в моче выше в группе с низкой ДБГ, что авторы интерпретировали как показатель сниженной активности НА — систем вследствие того, что МОФЭГ в первую очередь является индексом внутринейронального метаболизма, а сумма НА+НМ+ВМК отражает экстранейрональный метаболизм НА, который показывает скорость выброса НА и, соответственно, функциональный уровень НА систем. Однако последнее утверждение в свете многочисленных имеющихся данных вызывает большое сомнение (Kopin, 1985). Подводя итог этой части обзора, можно сделать по крайней мере два убедительных, как нам представляется вывода. Первый — функциональное состояние КА — систем при аффективных расстройствах безусловно нарушено по сравнению с нормальным состоянием и второй — эти изменения имеют прямое отношение к патогенезу депрессий. Противоречивость литературных результатов в значительной степени может объясняться разнообразием клинических вариантов расстройств, различными клиническими подходами авторов, наличием сопутствующих соматических заболеваний, различиями в возрасте и поле больных, длительностью медикаментозной терапии больных и многими другими факторами. Учитывая чрезвычайную полифункциональность КА — систем, следовало ожидать, что картина перестроек функциональных особенностей адаптивных механизмов будет иметь крайне гетерогенный характер, тем более, что неясны взаимоотношения между центральным и периферическим обменом КА. С этой точки зрения было бы крайне желательно провести комплексное исследование КА — систем метаболизма при аффективных нарушениях с одновременным определением широкого числа показателей метаболизма КА в разных биологических жидкостях. К сожалению, практически все работы, выполненные до сих пор, ограничиваются только определенными показателями, а целостная картина строится только на основе разнородных исследований. Второй момент, который можно отметить при анализе имеющегося материала, касается конечной цели исследований. В основном авторы ограничиваются вопросом: «имеются или нет объективные нарушения катехоламиновой или другой медиации при аффективных расстройствах»? Видимо, на данном этапе исследований, учитывая крайнюю противоречивость результатов, оправдано стремление авторов в первую очередь найти клинические основания для выделения вариантов депрессий с различными биохимическими детерминантами, подобрать маркеры и прогностические показатели к отдельным аффективным нарушениям. Пока, несмотря на многочисленные попытки, большими успехами этот путь не увенчался, хотя и были выделены разнородные группы депрессивных пациентов с различными сочетаниями параметров некоторых нейромедиаторных систем, разными клиническими особенностями и терапевтическими подходами (van Praag H.M. et al., 1990). Одной из попыток преодолеть противоречивость результатов явился подход, сформулированный van Praag H.M. (1990). Отказ от синдромологического подхода оказал плодотворное воздействие на развитие знаний о патохимии депрессий. Так, по мнению van Praag H.M., недостаточность индоламиновой медиации в первую очередь обусловливает развитие аффективных расстройств, центральная дофаминергическая система связана с двигательными нарушениями в рамках депрессий и других психических заболеваний, а функциональная патология норадренергических механизмов имеет отношение к проявлению симптомов ангедонии. Не вызывает никакого сомнения, что различные синдромологические проявления в рамках психических расстройств имеют под собой нейрохимическую основу в виде преимущественного поражения той или иной медиаторной системы и с этой точки зрения синдромологическая концепция van Praag H.M. хорошо объясняет многие известные экспериментальные факты. Вместе с тем трудно согласиться с утверждением о том, что, например, норадреналиновая система ответствена в первую очередь за проявления черт ангедонии в структуре различных психопатологических расстройств. Норадренергические нервные пути являются универсальными регуляторными структурами головного мозга (Foote, 1986) и клинические признаки недостаточности норадреналиновых структур должны быть значительно разнообразнее того, о чем говорят van Praag H.M. et al. (1990). С этой точки зрения, например, вызывает интерес участие катехоламиновых систем в патогенезе обсессивно-компульсивных расстройств и так называемых расстройств контроля импульса. Хотя роль серотониновых систем в патогенезе расстройств контроля импульса является общепризнанной, нельзя не согласится с тем, что центральные катехоламиновые механизмы не могут оставаться индифферентными в процессах формирования и развития этого типа психических расстройств. Это связано с тесным взаимодействием индоламиновых и катехоламиновых систем на структурно-функциональном и биохимическом уровнях, а также со схожестью нейрофизиологических особенностей серотониновых и норадреналиновых тормозных механизмов головного мозга человека. Эти соображения обусловили появления ряда работ, посвященных изучению взаимосвязи метаболизма катехоламинов с обсессивно-компульсивными расстройствами. Siever et al. (1983) и Rasmussen et al.(1987) обнаружили увеличение содержания НА и МОФЭГ в крови больных с обсессивно-компульсивными расстройствами (ОКР) по сравнению с контрольными показателями. Очевидно, что эти результаты напоминают данные обследования больных с депрессиями. Вместе с тем уровень МОФЭГ и ГВК в ликворе у больных с ОКР и содержание НА и А в плазме крови у подростков с ОКР не отличались от нормы. Не нашли отличий в содержании НА, А и основных метаболитов НА и ДА МОФЭГ и ГВК в плазме крови между ОКР и нормой Benkelfat et al.(1991). 7.1.4. Особенности обмена серотонина при психических нарушенияхСеротонин (5-окситриптамин) — один из основных медиаторов центральной нервной системы, выполняющий также важные гормональные функции на периферии организма. Серотонинергические синаптические окончания широко распространены практически во всех отделах головного мозга, серотониновые нейропроводящие пучки модулируют и регулируют функциональное состояние множества нейрональных систем. Гормональные функции широкого спектра действия серотонина на периферии включают вазоконстрикторные свойства, стимуляцию гладкой мускулатуры различных внутренних органов и ряд других функций. Нужно отметить, что все стадии биосинтеза и катаболизма индоламинов, физиологические особенности от образования и выброса до обратного захвата, этого типа нейромедиаторов чрезвычайно схожи с аналогичными этапами метаболизма, типичными для катехоламиновых систем. Отличительными чертами серотониновых нейропроводящих структур головного мозга являются: прежде всего локализация тел нейронов, использующих индоламиновые медиаторные возможности в ограниченном числе ядер, располагающихся главным образом в строго определенном регионе головного мозга. Основными ядрами, содержащими около 80 % серотонина головного мозга являются так называемые ядра шва среднего мозга. Порядка 10 ядер содержат тела индоламинергических нейронов, которые иннервируют практически все отделы мозга — кору больших полушарий, подкорковые ядра (включая стриатум и компоненты лимбической системы), гипоталамические образования, мозжечок, ствол головного мозга, а также спинной мозг (Azmitia, 1978; Azmitia, Gannon, 1986, Topel, 1985, Cooper et al., 1986). Аксоны этих нейронов оканчиваются практически во всех отделах мозга, формируя главным образом восходящие и нисходящие проводящие пучки (Azmitia, 1978; Tork, 1990). Подобная архитектура нейронной сети свидетельствует о том, что индоламиновые механизмы, подобно норадренергическим, участвуют практически во всех типах интегративных процессов головного мозга, и их роль сводится, скорее, к модуляторным и интегрирующим воздействиям на более специфические нейрональные системы (Rogeness et al., 1992; Spoont M.R., 1992). Не вдаваясь в подробный нейроанатомический анализ восходящих и нисходящих проводящих путей, формируемых серотонинергическими ядрами среднего мозга, следует отметить универсальность и всеохватность этой сети, что означает важность серотонинергической иннервации для синхронной работы систем головного мозга. Интересно, что некоторые проводящие пути отличаются филогенетической специфичностью — так, кортикальный тракт, терминали которого оканчиваются на нейронах коры больших полушарий, в гораздо выраженней степени развит у приматов по сравнению с грызунами (Azmitia, Gannon, 1986). Учитывая универсальную роль серотониновых структур, очевидно, что серотонинергические синаптические образования принимают участие в формировании и окончательном оформлении множества поведенческих и эмоциональных проявлений (Azmitia, 1987; Spoont M.R., 1992). Так, серотониновые механизмы контролируют формирование двигательных актов, систем положительного подкрепления, играют заметную роль в пищевом, половом, исследовательском поведении, участвуют в формировании аффективных компонентов поведенческих актов, определяют становление и поддержание суточных и циркадианных ритмов физиологических процессов, осуществляют температурную регуляцию организма. Нейрохимические механизмы сна в значительной степени реализуются на базе серотониновых структур. Общепризнано, что серотонин является основным химическим медиаторным соединением, контролирующим агрессивное поведение (Oliver, Mos, 1992) и выраженность тревожных проявлений на фоне меняющихся условий внешнего окружения (van Praag H.M. et al.,1990). Очевидно, что перечень функциональных возможностей центральной нервной системы, в реализации которых принимает участие серотонин, далеко не полон. Это вполне естественно, учитывая анатомические особенности серотонинергической иннервации мозговых структур (Spoont M.R., 1992). Ряд авторов полагает, что уровень серотонина (5-ОТ) в плазме крови может предоставить достоверную информацию относительно уровня активного трансмиттера в синаптической щели центральных синапсов (Celada P. et al., 1990). Как бы фантастично ни выглядело это предположение, ряд экспериментальных фактов свидетельствует о реальности подобной возможности. Так, пулы серотонина и его основного метаболита 5-оксииндолуксусной кислоты (5-ОИУК) в плазме являются независимой популяцией, поведение которой зачастую не совпадает с динамикой перестроек содержания 5-ОТ в тромбоцитах (Artigas F. et al., 1989). Об этом свидетельствует отсутствие корреляции между уровнем серотонина в плазме и содержанием этого вещества в цельной крови; если бы экстрацеллюлярный пул серотонина плазмы был обусловлен исключительно разрушением тромбоцитарных клеток, упомянутая корреляция обязательно бы присутствовала (Ortiz et al., 1988). Различные воздействия включая введение ингибиторов моноаминоксидазы (МАО-А) и ингибиторов синтеза серотонина, приводят к снижению концентрации серотонина и 5-ОИУК в плазме, но не меняют уровень 5-ОТ в тромбоцитах (Artigas F. et al., 1985; Ortiz et al., 1988, Celada P. et al., 1990). Длительная терапия ингибиторами обратного захвата серотонина приводит к противоположному эффекту: снижается содержание медиатора в тромбоцитарных клетках, а плазменный пул остается неизменным (Sarrias M.J. et al., 1987). Противоположные эффекты были получены также при терапии больных с депрессиями солями лития: увеличение уровня серотонина в плазме, отсутствие эффекта в тромбоцитах (Artigas F. et al., 1989). Эти, а также другие результаты дают возможность полагать, что серотониновые молекулы плазмы крови составляют популяцию с высокой скоростью кругооборота молекул, быстро реагирующую на введение агентов, воздействующих на метаболизм серотонина, в то время как пул серотонина в тромбоцитах является более стабильной популяцией, меняющейся под влиянием более длительных воздействий. Вышеприведенные факты дали возможность Meltzer (1989), Ortiz et al. (1988, 1991), Artigas F. et al. (1989), Celada P. et al. (1990) высказать мнение о схожести динамических взаимоотношений между тромбоцитами и окружающей плазмой, с одной стороны, и равновесием, имеющим место между пресинаптическими окончаниями и экстрацеллюлярной жидкостью синаптической щели, с другой. Проблема патогенетических изменений индоламиновых механизмов при депрессивных расстройствах к настоящему времени отнюдь не кажется разрешенной. Общая картина в чем-то напоминает ситуацию с катехоламиновой системой. Огромное количество исследований, проведенное в последние 30 с лишним лет, убеждают лишь в одном — серотониновые системы играют важную роль в патогенезе аффективных расстройств. Однако противоречий в современных взглядах на понимание механизмов, ведущих к проявлению психических нарушений едва ли не больше, чем в случае с катехоламиновыми механизмами (Дроздов А.З., 1997). К настоящему времени уже прочно сложилось мнение о необязательности уменьшения содержания 5 — оксииндолоуксусная кислота 5ОИУК в спинномозговой жидкости (СМЖ) при депрессивных расстройствах (Caldecott-Hazard et al., 1991; Reddy et al., 1992; Leonard, 1995). Также очевидно, что данный факт не следует связывать только с методологическими трудностями, обусловленными такими неспецифическими факторами, как возможная зависимость показателей индоламинового обмена от возраста, пола, веса, циркадианных и сезонных ритмов (Asberg et al., 1984) или тем, что люмбарная пункция может предоставлять биологический материал, отражающий главным образом спинальный метаболизм серотонина, даже и при условии наличия достоверной корреляции между цистернальным и спинальным уровнями 5ОИУК (Degrell, Nagy, 1990). В конечном итоге ненадежность такого признака депрессивных расстройств, как дефицит количества основного продукта метаболизма серотонина в СМЖ больных, обусловлена, видимо, не методическими артефактами, а скорее несоответствием нозологического уровня психопатологического явления и более частным характером биологической дисфункции как отражения определенных особенностей генетического аппарата организма. В этой связи заслуживает рассмотрения ряд доказательств того, что сниженная концентрация 5ОИУК в ликворе является весьма частым спутником таких психических особенностей, как суицидальные тенденции и импульсивность (van Praag H.M., 1986; Mann et al., 1989, 1992а; Cremniter et al., 1994; Mehlman et al.,1994), повышенные агрессивность (Virkkunen M. et al., 1987; Mann et al., 1989; McBride et al., 1990; Higley et al., 1992; Mehlman et al., 1994) и тревожность (Rydin et al., 1982; van Praag H.M. et al., 1990). Нужно отметить, впрочем, что увеличение содержания 5ОИУК в СМЖ по сравнению с контрольными значениями иногда отмечается при обсессивно-компульсивных расстройствах (Insel T.R. et al., 1985; Leonard, 1989). Имеются, правда, свидетельства об отсутствии среднегрупповых различий между больными с обсессиями и здоровыми людьми по данному параметру, но при существовании достоверной отрицательной корреляционной связи между уровнем 5ОИУК ликвора и тяжестью некоторых обсессивных проявлений (Thoren et al., 1980; Swedo S.E., et al., 1992), а также достоверной положительной зависимости между 5ОИУК ликвора и клиническими симптомами выздоровления пациентов (Swedo S.E. et al., 1992). Содержание самого серотонина и его основного метаболита было снижено в разных отделах головного мозга у больных с синдромом Жиля де ля Туретта (Anderson et al., 1992), заболеванием, симптомы которого могут являться одним из проявлений обсессивно-компульсивных расстройств (Robertson et al., 1988; Swedo S.E. et al., 1992). Таким образом, несмотря на ряд противоречивых мнений, обсессивно-компульсивные расстройства также сопровождаются изменением центральных индоламиновых функций по типу их угнетения. Комплекс имеющихся данных позволяет сделать предположение о непосредственном отношении сниженного функционального состояния индоламиновых систем ЦНС к проявлению компульсивных и импульсивных нарушений поведения. Подобная интерпретация хорошо вписывается в схему тормозной модулирующей структурно-функциональной организации серотониновых механизмов головного мозга (Spoont M.R., 1992). Широко распространенные в современной психиатрии нейроэндокринные тесты с очевидностью демонстрируют выраженные нарушения индоламиновой регуляции центральных процессов при ряде психопатологических состояний — депрессиях, обсессивно-компульсивных расстройствах, психопатиях, агрессивных проявлениях, при этом авторы с осторожностью рассматривают причины этих дисфункций и не рискуют оценивать даже качественное состояние (гипо- или гиперфункция) серотониновой медиации при перечисленных психических нарушениях, хотя низкие нейрохимические отклики на серотониновые агонисты, наверное, можно интерпретировать как функциональную недостаточность центральных серотониновых систем. Одной из основных причин подобного состояния проблемы является чрезмерная сложность изучения функционального статуса рецепторного аппарата моноаминовых систем мозга. Существование множества подтипов рецепторов, различная локализация (пре- и постсинаптическая), разнонаправленность физиологического действия при связывании агонистов и антагонистов, наличие множества эндогенных лигандов смешанного типа действия, наличие рецепторов для разных нейроактивных веществ на одних и тех же нейронах, различное состояние однотипного рецепторного аппарата в различных отделах головного мозга, также как ряд иных факторов, все это сильно осложняет анализ клинико-биохимических данных. Одним из психопатологических проявлений, при котором обнаруживается увеличение содержания серотонина в тромбоцитах или цельной крови, является повышенная агрессивность и импульсивность (Raleigh et al., 1984; Pliszka et al., 1988; Mann et al.,1992). Учитывая снижение уровня 5-ОИУК в ликворе при агрессивности, особенно импульсивной агрессии (Brown S.L. et al.,1982; Virkkunen M. et al.,1987,1989а,б), Mann et al.(1992) делают предположение о возможном увеличении уровня серотонина в тромбоцитах как о проявлении наследственного феномена усиления процесса обратного захвата моноаминов (или снижения их высвобождения) в пресинаптических терминалях ЦНС и в тромбоцитах, что приводит к недостаточности моноаминовых механизмов. 7.1.5. Серотонин и половая дифференцировка мозгаПоловой диморфизм отражается на нескольких уровнях: генетическом, гонадном, морфологическом, гормональном и церебральном. Исходя из концепции о многоуровневой организации пола (Васильченко Г.С., 1990; Кон И.С., 1989) можно выделить различные нарушения его формирования на разных уровнях, что, однако, трудно представить как локальное явление, не затрагивающее других звеньев такой сложной системы, каковой является пол. По современным представлениям, пол человека представляет собой многоуровневую иерархическую систему, складывающуюся из параллельно протекающих, но во многом взаимно обусловленных процессов биологического и социального развития индивидуума. В основе формирования системы пола лежит процесс половой дифференцировки мозга (ПДМ), начинающийся в раннем эмбриональном периоде и продолжающийся вплоть до полного созревания. В системе формирования пола одну из ведущих ролей играют нейромедиаторные системы. Ряд авторов склонны предполагать, что нейромедиаторы и нейромодуляторы выполняют в организме роль универсального кода, отвечающего за всю психическую деятельность и соматопсихическое развитие в целом (Белкин А.И., Ракитов А.И., 1989). Непосредственную регуляцию в системе гипофиз-гонадотропины-гонады, возможно, выполняют и простогландины (Вундер П.А. 1980). О возможности участия серотонина в дифференцировке полового поведения свидетельствует снижение частоты маунтинга и выраженные реакции по женскому типу у самцов, неонатально получавших p-хлорфенилаланин, являющийся ингибитором синтеза серотонина. Напротив, при введении ингибиторов моноаминоксидазы (МАО), которые способствуют накоплению катехоламинов и серотонина в нервной ткани, у самок отмечалась маскулинизация поведения. Таким образом, серотонин является своего рода детерминирующим фактором мужского полового поведения. Основным медиатором, опосредующим влияние андрогенов на ПДМ, является норадреналин (Резников А.Г., 1982). Вмешательство в ПДМ в критические периоды (неонатальная андрогенизация самок) можно идентифицировать морфологически (организация аркуатных ядер гипоталамуса по мужскому типу (Matsumota A., Yasumasa A., 1981) Половой диморфизм миндалевидного комплекса (МК), связан с дофамин- и НА-путями. (Karakiulakis G., et al. 1978.). Это подтверждается данными о существовании разного количества дофаминергических и серотонинергических рецепторов в МК у мужчин и женщин. В последнее время накоплено достаточно фактов, свидетельствующих в пользу влияния изменений в серотониновой системе непосредственно на сексуальное поведение (Spoont M.R., 1992). Так, в результате ряда исследований было установлено, что снижение содержания серотонина в головном мозге приводит к повышению половой активности животных. Исходя из имеющихся фактов, исследователи пришли к выводу, что серотонинергические механизмы оказывают ингибирующее влияние на половое поведение животных. В пользу данного вывода свидетельствовали так же факты, как усиление гомосексуального поведения самцов после снижения уровня серотонина в мозге на 80 %, исчезновение половой активности после введения 5-окситриптофана (предшественника серотонина в цепи биосинтеза) и другие (Gessa G.L., 1978) . Снижение уровня серотонина в головном мозге сопровождается усилением полового влечения у всех животных, однако на внешние проявления гиперсексуальности определенный отпечаток накладывает «социальное положение» животного, его место в иерархии (Spoont M.R., 1992). В соответствии с современными представлениями, парафилии представляют собой нарушение психосексуальных ориентаций, являющихся завершающим этапом психосексуального онтогенеза. В свете целостного понимания психосексуального развития как единого непрерывного процесса, первым этапом которого является формирование полового самосознания и полоролевого поведения, лица с парафилиями оказываются в группе риска с различными вариантами нарушения половой идентичности. Последние, с одной стороны, создают комплекс клинико-психологических предиспозиций для развитий аномалий сексуального влечения, с другой стороны указывают направление поиска клинико-патогенетических звеньев данных расстройств. У обследованных пациентов мы определяли содержание свободных и конъюгированных форм норадреналина, адреналина, дофамина в плазме крови и суточной моче; содержание предшественника катехоламинов в цепи биосинтеза — диоксифенилаланина (ДОФА) и метаболита дофамина диоксифенилуксусной кислоты в плазме крови и в суточной моче; концентрацию серотонина в тромбоцитах и в плазме крови; скорость захвата меченого серотонина тромбоцитами. Определение содержания всех исследуемых веществ проводили при помощи различных методик, в частности используя высокоэффективную жидкостную хроматографию с электрохимическим детектором (Коган Б.М., Дроздов А.З., Маньковская И.В., Филатова Т.С. 1994 — в печати., Андрианова Е.П., Дроздов А.З., Жуковская Е.Д., Куканова М.М., Морозова Г.В., Наумович А.О., Коган Б.М. 1994). Основные параметры (K 4m 0 и V 4max 0)»захвата» серотонина тромбоцитами определяли по методу (Брусов О.С. и соавт. 1988) с небольшими модификациями. Статистическую обработку результатов проводили используя критерий Стьюдента. 7.2. Особенности метаболизма нейромедиаторов при парафилиях7.2.1. Конституцинально-дизонтогенетические предиспозиции сексуальных нарушений и нейромедиаторыСреди биологических факторов, влияющих на патогенез парафилий, первостепенное значение имеют аномалии тех нейрофизиологических и биохимических процессов, которые сопряжены с составляющими полового диморфизма. Нарушения биологических предиспозиций половой дифференцировки и определяемые ими отклонения процессов половой идентичности и психосексуальной ориентации тем или иным образом отражаются в феноменологических проявлениях различных форм аномального сексуального поведения. В связи с этим обнаружение мотивационных патопсихологических и психопатологических паттернов парафилий возможно в тех сферах личности, которые являются производными от полоролевых стереотипов и других конструкт маскулинности/фемининности. При понимании основной детерминирующей роли аномалий полового диморфизма становится возможным проведение и клинико-биохимических сопоставлений.Удалить? Прежние исследования касались прежде всего изучения гормонального гомеостаза, поскольку именно гормональный фактор у мужчин оказывается решающим в окончательном преобразовании механизмов маскулинизации и дефеминизации. Однако ясно что весь биохимический комплекс, составляющий нейроэндокринную предрасположенность к полоспецифическому поведению, не ограничивается только гормональным гомеостазом, но включает все остальные звенья нейрогуморальной регуляции. В частности, моноаминовые нейромедиаторные системы контролируют функционирование некоторых эндокринных систем, регулирующих половое поведение [24], а также непосредственно участвуют в пренатальных процессах половой дифференцировки мозга [4,10,16]. Таким образом, психосексуальное развитие оказывается неразрывно связанным в первую очередь с конституционально-дизонтогенетическими предиспозициями, представляющими собой комплекс морфосоматических, нейрофизиологических и биохимических оснований индивидуального развития. В связи с вышеизложенным нам представлялось важным проведение исследования состояния основных звеньев моноаминергической нейромедиации у лиц с аномальным сексуальным поведением, отличающихся по ряду конституциональных и поведенческих особенностей. При выборе конституциональных параметров мы исходили из того, что в сексологической практике наиболее широкое распространение получила методика морфограмм предложенная J.Decourt и J. Doumic (1950) По этой методике определяется пять параметров: 1. Рост 2. Высота ноги (от верхнего края большого вертела до площади опоры) 3. Окружность грудной клетки (в промежуточной позиции между вдохом и выдохом) 4. Бигумеральная (межплечевая) дистанция 5. Расстояние между spina aliaka anterior superior На основании произведенных измерений J.Decourt и J. Doumic выделили нормальные половые биотипы — мужской и женский. Так же в сексологическом обследовании был особо выделен трохантерный индекс (Т.И.), который отражает один из описанных параметров — отношение длины ноги к росту. Теоретическим основанием для выделения Т.И. послужило учение о периодичности роста человеческого организма, в частности положение высказанное Н.П.Шаповаленковым (1927): «В препубертатном периоде рост увеличивается удлинением ног, в следующим непосредственно за ним пубертатном — удлинением торса.» Из работ В.В. Бунак (1954, 1960) можно сделать вывод, имеющий особое значение для сексологической антропометрии: «Диаэпифизарная зона, как правило, не реагирует на механические стимулы, и относительная длина сегментов конечностей составляет существенный видовой признак, который едва ли может быть поставлен в зависимость от случайных колебаний механической нагрузки скелета у отдельных индивидуумов». Трохантерный индекс является признаком специфическим, и его отклонения свидетельствуют о наличии раннего нарушения пубертатного развития. В нашем исследовании использовались следующие параметры оценки трохантерного индекса: 1. Т.И. меньше или равен 1,91 2. Т.И. от 1,91 до 1,98 3. Т.И. равен или больше 1,99, ; что соответствует слабой, средней и сильной половой конституции. Так же конституция оценивалась с использованием индекса R.E., который вычислялся по формуле: R.E.= рост умноженный на сто и деленный на ширину грудной клетки, умноженную на шесть. Существует три типа конституции: 1. астеническая R.E.> 105 2. нормостеническая R.E. 95,2- 104,3 3. гиперстеническая R.E. Андрогиния определялась с использованием индекса Таннера, который вычислялся по формуле: И.Т. = биакромиальный размер умноженный на три минус бикристальный размер. В настоящее время описано три типа 1. андромофный И.Т.> 93,5 2. мезоморфный И.Т. 87,0 — 93,5 3. гинеморфный И.Т. Нейрохимическое исследование показало, что традиционная для сексологической практики группировка испытуемых по сходным конституционально-анатомическим параметрам не позволяет обнаружить стойкие клинико-биохимические корреляции. Суточная экскреция катехоламинов и их метаболитов у больных, представляющих группы лиц с различным трохантерным индексом, конституциональными и андрогинными особенностями, практически не различалась. Подобная картина отмечается также и по показателям моноаминового метаболизма в крови. Тем не менее можно отметить ряд закономерностей, выявленных в ходе выполнения настоящего фрагмента работы. Так, заслуживает упоминания некоторое снижение концентрации связанной формы ДА в андрогиной группе 3 по сравнению с группой 2 и контрольной группой (табл. 7.1). Достаточно трудно интерпретировать подобную закономерность без привлечения результатов исследования иных физиологических систем. Тем не менее общий смысл снижения уровня конъюгированных форм ДА — катехоламина, присутствующего в плазме почти исключительно в конъюгированной форме, — может заключаться в уменьшении интенсивности инактивационного метаболического конъюгационного пути. Известно, что общие тенденции метаболизма активных моноаминовых молекул у женщин заключаются в увеличении скорости метаболического разрушения катехоламинов, что объясняется увеличением активности основного энзима метаболизма КА — МАО по сравнению с мужчинами. Согласно полученным результатам, для конъюгационных процессов характерна противоположная тенденция, что проявляется в признаках ослабления интенсивности конъюгационных процессов (например, снижение уровня конъюгированного ДА плазмы крови) по мере усиления гинекоморфных конституциональных особенностей. Данный аспект физиологии медиаторных систем практически не изучен и на настоящем этапе исследований можно констатировать, что сам по себе факт присутствия клинических признаков парафилий может не иметь отношения к среднегрупповым различиям по катехоламиновым параметрам между лицами с отличающимися конституциональными характеристиками. Таблица 7.1. Содержание свободных и конъюгированных форм катехоламинов, а также ДОФА и ДОФУК в крови испытуемых с парафилиями
* — р Также заслуживают внимания и достоверные различия между по уровню тромбоцитарного серотонина, которые непросто поддаются адекватной интерпретации. Сам по себе уровень различий свидетельствует о неслучайности зафиксированных закономерностей. Основное отличие сводится к резкому увеличению содержания внутриклеточного серотонина в группе c астенической конституцией по сравнению с испытуемыми с нормостенической и гиперстенической конституцией (табл. 7.2). Уровень серотонина тромбоцитов в последних двух группах не отличается от контрольных показателей (табл. 7.2). Увеличение концентрации серотонина в тромбоцитарных клетках характерно для больных с тревожно-депрессивными и паническими расстройствами (Дроздов А.З., 1996). Анализ результатов наших исследований свидетельствует о том, что подобная картина динамики распределения различных пулов серотонина крови отражает не столько определенное клиническое состояние, сколько генетически обусловленное состояние индоламиновых медиаторных систем, предрасполагающее к проявлению определенных характерологических особенностей. Очевидно, что вне зависимости от клинических проявлений астеническая конституция очерчивает группу риска развития тревожных и депрессивных состояний по сравнению с нормостенической и гиперстенической конституцией, в связи с чем высокие цифры внутриклеточного серотонина в астенической группе можно объяснить в свете известных экспериментальных фактов (Дроздов А.З., 1996). В данном случае результаты вновь прямо не указывают на специфическую связь метаболизма моноаминов с симптомами собственно парафилий. Таблица 7.2. Уровень серотонина в тромбоцитах и в плазме крови у испытуемых с парафилиями
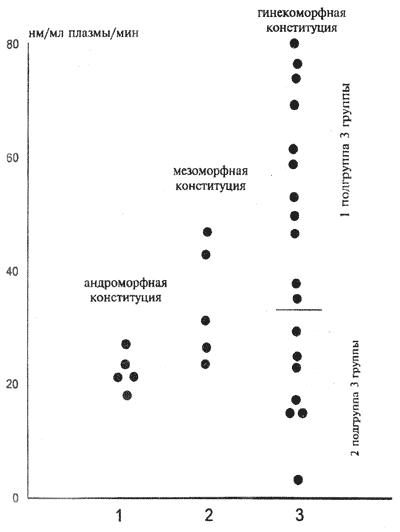
* — р ** — р При изучении клинико-биохимических взаимосвязей у лиц с парафилиями особый интерес вызывает исследование активности дофамин-b-гидроксилазы — фермента, активность которого находится под генетическим контролем и стабильна на протяжении жизни человека. ДБГ является одним из лимитирующих ферментов биосинтеза норадреналина. Он катализирует последнюю стадию биосинтеза норадреналина (НА) — гидроксилирование дофамина с образованием активного нейромедиатора. Фермент секретируется в синаптическую щель из терминалей нейронов симпатической системы вместе с НА, после чего попадает в кровоток. Согласно данным ряда исследователей, активность этого фермента может отражать функциональное состояние симпатической нервной системы. Кроме того, имеется прямая корреляционная зависимость между активностью ДБГ в крови, спинномозговой жидкости и мозге, что свидетельствует о возможности оценки скорости последней лимитирующей реакции биосинтеза НА в нейронах мозга по степени активности периферической ДБГ. Как указывалось выше, в результате антропометрического обследования всех испытуемых образовались три группы, соответствующие различным конституциональным типам полового диморфизма. Первую группу больных составили андроморфы, вторую — мезоморфы и третью — гинекоморфы. Значительное преобладание испытуемых с парафилиями с гинекоморфной конституцией по сравнению с индивидуумами других конституциональных типов может быть оценено только при дальнейшем сопоставлении данных с закономерностями подобного распределения в общей популяции. Тем не менее даже при самом осторожном подходе к интерпретации результатов выявление преобладания гинекоморфного телосложения среди лиц с парафилиями, по крайней мере, может свидетельствовать о своеобразном характере их дизонтогенеза, при котором задержка развития сопровождается сохранением фемининных пропорций телосложения. Последние могут являться одним из биологических признаков изменения процессов, связанных с половой дифференцировкой. Первую группу составили испытуемые с наиболее яркими по сравнению с другими обследованными проявлениями органического поражения головного мозга с преобладанием гипоталамической патологии. Клинически это выражалось в эндокринной дисфункции с задержкой пубертатного развития, склонности к диэнцефальным пароксизмам и перманентным вегетососудистым расстройствам. Патология сферы влечений носила диффузный характер и включала в себя и иные виды, кроме сексуального (так, в двух случаях имело место клептоманическое поведение в подростковом периоде). Окончательное формирование аномального сексуального влечения, во всех случаях имевшего агрессивно-садистическую структуру независимо от характеристик объекта влечения (в двух случаях — гомосексуальный, в остальных — гетеросексуальный), происходило по миновании пубертатного периода. Во вторую группу вошли лица, у которых имелись четкие признаки психопатической структуры личности с преобладанием особенностей, характерных для круга тормозимых. Лишь один обследованный обнаруживал агрессивно-садистические тенденции, тогда как в остальных случаях имели место гомосексуальное, эксгибиционистское и фетишистское поведение. Третья, самая многочисленная группа, оказалась и наиболее клинически разнородной. В ходе дальнейшего исследования в ней были выделены две подгруппы, описание которых приводится ниже. В целом можно сказать, что все выделенные группы характеризовались различным вкладом участвующих в формировании аномального сексуального поведения предиспонирующих факторов — конституционально-дизонтогенетических, органических, патохарактерологических и психопатологических. Распределение активности ДБГ в плазме крови в исследованных группах представлено в таблице 7.3 и на рисунке 7.1. Наиболее низкая активность фермента обнаружена в группе больных с андроморфным типом конституции, наиболее высокая — у гинекоморфных больных. Известно, что разброс нормальных значений очень широк (примерно 0,1 — 110 нМ/мл/мин). Если в группах пациентов с мезо- и андроморфным телосложением показатели активности ДБГ в крови носили однонаправленный характер, то наиболее многочисленная группа больных с гинекоморфным телосложением оказалась внутренне неоднородной и включила в себя две подгруппы с различной активностью ДБГ (рис. 7.1). Такой характер выявленных изменений позволил провести клиническое сопоставление образовавшихся подгрупп. Таблица 7.3. Активность дофамин-и-гидроксилазы в плазме крови у лиц с парафилиями
* — р *** — р + — р Выявилось несколько психопатологических и патосексологических критериев, указывающих на явное клиническое своеобразие данных подгрупп. В первую очередь они характеризовались различием феноменологических проявлений сексуального поведения. Если во 2 подгруппе в основном наблюдалось гетеросексуальное насилие, то в 1 подгруппе преобладали гомопедофильные действия или малодифференцированный выбор объекта сексуального влечения. Данные особенности аномального сексуального поведения базировались на более глубоких изменениях полоролевых установок, которые у лиц, составивших 1 подгруппу, характеризовались четкой тенденцией к трансформации полоролевого поведения. Имелись и клинико-динамические отличия, так как в первой подгруппе отмечалось прогрессирующее течение сексуального влечения, садистические элементы которого выявлялись по мере постепенного преобразования первоначально инвертированных пассивных тенденций, что позволяло рассматривать собственно агрессивные проявления в качестве вторичных образований, формирующихся в результате определенных защитных механизмов. Это определяло и отличие в экспертных подходах к обеим подгруппах. Во 2 подгруппе в основном учитывались традиционные критерии оценки невменяемости, причем в подавляющем количестве наблюдений имелись достаточные клинические основания для вынесения заключения об экскульпации. В 1 же подгруппе, при отсутствии убедительных данных о тяжести собственно психических расстройств, экспертные акценты смещались в сторону аномального сексуального влечения, которое само по себе приобретало патологические признаки и обретало юридическое значение.
Анализ биохимической части результатов показал, что формирование морфоконституциональных детерминант, изучаемых в настоящей работе, не зависит от состояния одного из лимитирующих ферментов биосинтеза НА. Так, в группе с мезоморфной конституцией встречаются больные и с более низкой активностью фермента, чем у людей с андроморфной, и с более высокой, чем в группе с мезоморфной конституцией (рис.7.1), хотя средняя активность в последней группе выше среднего уровня и достоверно превышает низкую среднюю активность ДБГ у лиц с андроморфной конституцией. Достаточно сложно установить жесткую взаимосвязь между процессами детерминирования конституциональных проявлений и наследственно определенной активностью ДБГ. Тем не менее локализация лиц с высокой активностью фермента только в гинекоморфной группе испытуемых (в отличие от андроморфной группы) свидетельствует о возможной сцепленности некоторых конституциональных особенностей с активностью важнейшего фермента анаболизма — норадреналина. Клинический анализ показал, что две подгруппы больных в рамках группы с гинекоморфным сложением, выделенные на основе различий в активности ДБГ, оказались различными и в клиническом плане. Клиническая дифференцировка подгрупп больных с высокой и низкой активностью фермента дает основание полагать, что особенности проявляемых больными аномальных сексуальных реакций зависят от врожденных свойств катехоламиновых нервных систем. Многочисленные данные свидетельствуют о том, что состояние НА-ергических систем мозга, косвенным индикатором которого является активность ДБГ плазмы крови, связано с выраженностью определенных личностных особенностей, причем низкая активность ДБГ может приводить к проявлению черт характерологической неустойчивости, особенно заметных в условиях неблагоприятного внешнего окружения. Низкая активность фермента обнаружена у лиц, склонных к развитию реактивных депрессивных и шизофреноподобных состояний, у больных алкоголизмом, дающих психотические реакции на фоне лечения дисульфирамом. Наиболее низкая активность ДБГ в крови отмечается у больных алкоголизмом с преморбидными астеническими и истеровозбудимыми чертами характера. Обнаружена достоверная отрицательная корреляционная зависимость между активностью ДБГ в ликворе и выраженностью таких шкал MMPI как ипохондрия, депрессия истерия, психопатия, психастения и социальная интраверсия. Таким образом, у лиц второй подгруппы имеющиеся психические расстройства являются скорее проявлениями общей неустойчивости высшей нервной деятельности, характерной для индивидов с низкой активностью ДБГ, на фоне относительно сохранной структуры сексуальных влечений. Норадреналиновая система у больных первой гинекоморфной подгруппы по сравнению со второй отличается более надежным функциональным состоянием, что связано с достаточными резервными возможностями биосинтеза медиатора. У этих больных при большей эмоциональной устойчивости и стеничности по сравнению с лицами второй подгруппы отмечаются более глубокие нарушения высших центров контроля сексуального поведения. Тенденция к низкой активности фермента во 2 подгруппе гинекоморфных пациентов аналогична распределению активности в группе больных с андроморфным конституциональным типом (табл.7.3). Интересно, что эти две группы пациентов объединяют общие клинические особенности психических нарушений, которые проявляются в преобладании дефицитарных расстройств психики либо за счет психоорганической патологии, либо за счет процессуально нарастающих негативных изменений личности. Сходным было и клиническое проявление аномального сексуального поведения, сразу находившего агрессивно-садистическую форму своего выражения, что может объясняться присущей этим группам больных слабостью тормозных норадреналиновых структур головного мозга. Таким образом, не вызывает сомнений факт, что наблюдаемые различия в клинической картине сексуальных расстройств в определенной степени связаны со структурой личностных и психопатологических особенностей пациентов, определяемых состоянием моноаминовых систем головного мозга. Вместе с тем мы не можем исключить и непосредственную связь формирования определенных симптомов аномального сексуального поведения с особенностями моноаминовых систем, контролирующими половые функции организма. 7.3.2. Нейромедиаторы и искажения половой идентичностиОбследованы 104 испытуемых, совершивших сексуальные правонарушения и проходивших судебно-психиатрическую экспертизу в ГНЦ С и СП им. В. П. Сербского в период с 1993 по 1995 гг. Преобладали (61 чел.) лица с органическим поражением головного мозга различной степени выраженности и различной этиологии (из них с парафилиями 36), среди лиц с органическим поражением головного мозга с эписиндромом 3 человека, из них с парафилиями 1. На втором месте по частоте (19 чел.) оказались лица с различными видами личностных расстройств (из них с парафилиями 13), на третьем месте (15 чел).больные с диагнозом шизофрении (из них с парафилиями 5), после них больные с олигофренией (5 чел), из них с парафилиями 2). Двое обсследованных признаны психически здоровыми. Лица, находившиеся в момент исследования в состоянии реактивного психоза (3 чел.) в дальнейшем были исключены из статистической обработки. Среди 56 больных с парафилиями у 10 диагностирован садизм, у 28 — педофилия и эфебофилия (гомосексуальная и гетеросексуальная), у 6 — полиморфный парафильный синдром, у 4 — эксгибиционизм, у 2 — некрофилия. Остальные 48 испытуемых не обнаруживали признаков парафилий, несмотря на противоправное сексуальное поведение. Для оценки половой идентичности лиц с сексуальными правонарушениями использовалась разработанная в лаборатории судебной сексологии центра, шкала типирования нарушений половой идентичности. В основу создания шкалы был положен онтогенетический принцип развития половой идентичности человека и принцип многоступенчатой иерархической системы пола (Васильченко Г.С., 1990). На основании экспертных оценок в шкалу был включен ряд психологических, клинических и поведенческих феноменов, отражающих становление половой идентичности индивида с детства до периода зрелости. Наличие нарушения половой идентичности проводилась путем вычисления общего интегративного показателя, позволяющего в целом оценить все обнаруженные у пациента признаки, свидетельствующие о нарушении половой идентичности. Необходимо указать, что констатация нарушения половой идентичности, определенного путем шкалирования, не сопоставима с клинико-нозологическими формами, фигурирующими в принятых в настоящее время классификациях (DSM-III-R, МКБ-10). На основании данного интегративного показателя, после типирования по шкале исследованных больных они были объеденены в четыре группы, в зависимости от выраженности искажения половой идентичности. В первую группу вошли лица, у которых после проведенного исследования не было выявлено нарушений половой идентичности (29 чел.), во вторую — больные с незначительно выраженным искажением половой идентичности (47 чел.), третью группу составили испытуемые с выраженным (23 чел.) и четвертую группу — с резко выраженным искажением половой идентичности (5 чел.). В контрольную группу включены 14 практически здоровых мужчин сопоставимого возраста. Ожидаемым оказалось нарастание количества лиц с парафилиями по мере усиления нарушений половой идентичности: в первой группе парафилии встречались в 11 случаях (37,93%), во второй — в 23 (48,93%), в третьей — в 17 (73,91%), в четвертой группе парафилии отмечались у всех испытуемых. Анализ результатов, полученных при изучении экскреции с мочой катехоламинов, демонстрирует значительные нарушения метаболизма этого класса нейромедиаторов у лиц с нарушениями половой идентичности. Обращает на себя внимание тот факт, что у представителей первой группы, т.е. на фоне ненарушенной половой идентичности фиксируется существенное снижение (по сравнению с нормативными значениями) уровня как свободных, так и сульфоконъюгированных форм норадреналина, адреналина и дофамина (табл.7.4). Резкое возрастание у этих испытуемых экскреции 3,4-диоксифенилаланина и нормальное содержание в моче 3,4-диоксифенилуксусной кислоты заставляют предполагать формирование у данного контингента своеобразного «блока» на стадии биосинтеза ДА, так как нет оснований думать об усилении катаболизма катехоламинов. Однако нам представляется, что традиционное сравнение со здоровыми людьми в данном случае является некорректным, так как нет данных о жесткой взаимосвязи между нарушением половой идентичности и состоянием определенных этапов метаболизма нейромедиаторов. Такой подход диктует проведение сравнения между пациентами с различными по выраженности нарушениями половой идентичности. Действительно, само наличие нарушения половой идентичности проводит к статистически достоверному увеличению экскреции с мочой свободных и сульфоконъюгированных форм норадреналина, адреналина и дофамина (табл.7.4). Интересно отметить, что рост количества катехоламинов идет параллельно нарастанию нарушений половой идентичности на фоне снижения содержания ДОФА и ДОФУК у представителей третьей группы (табл.7.4). Можно предполагать, что у испытуемых, отнесенных на основании клинического обследования к 3 группе, формируется комплекс патологических перестроек в метаболизме нейромедиаторов катехоламиновой природы, основными проявлениями которых можно считать смещение биохимического равновесия в сторону образования биологически активных нейромедиаторных молекул с их последующей сульфоконъюгацией, которая становится ведущим путем физиологической инактивации, т.к. снижение концентрации в моче ДОФУК — продукта окислительного дезаминирования дофамина — говорит о подавлении этого пути ферментативной деградации моноаминов. Если же учитывать точку зрения некоторых ученых, считающих, что конъюгированные формы катехоламинов в определенных условиях могут являться депо активных (свободных) молекул, то возникает еще большая уверенность в том, что выраженные нарушения половой идентичности сопровождаются существенной активацией и норадренергической и дофаминергической систем в организме. Таблица 7.4. Экскреция свободных и конъюгированных форм катехоламинов, свободных форм ДОФА и ДОФУК при парафилиях
* — р ** — р + — р $$ — р # — р ' — р Исследование показало, что группы, сформированные по принципу нарушения половой идентичности, с одной стороны, отличаются по периферическим параметрам индоламиновой системы друг от друга, а с другой — от контрольной группы (табл 7.5). Обращают на себя внимание следующие особенности групповых значений исследуемых показателей. Во-первых, очевидно, что между групповыми средними величинами имеются достоверные различия, которые касаются содержания серотонина в тромбоцитах и в плазме крови. К настоящему времени достаточно разработаны критерии использования тромбоцитарных клеток в качестве модельного объекта центральных пресинаптических окончаний (Stahl S.M., 1985), дающие возможность содержательной интерпретации обнаруженных феноменов. Во-вторых, общая тенденция изменения значений серотонина в плазме в группах с сексуальными расстройствами сводится к увеличению концентрации медиатора в плазме и уменьшению его содержания в тромбоцитарных клетках. Сами по себе эти факты свидетельствуют о возможном снижении скорости захвата серотонина тромбоцитами, причем уровень серотонина в тромбоцитарных клетках можно рассматривать в качестве интегративного маркера скорости захвата серотонина пресинаптическими окончаниями (Stahl S.M., 1985), а концентрация серотонина в плазме может отражать его уровень в синаптических пространствах мозга (Artigas F. et al., 1989; Celada P. et al., 1990). Так, средняя концентрация серотонина в плазме у больных 2 группы достоверно превышает контрольный уровень и показателе в 1 группе. Наиболее низкое содержание серотонина в тромбоцитах среди всех групп обнаружено в 1 группе, причем отличие имеет достоверный характер относительно контрольной группы (табл.7.5). Очевидно, что наиболее низкий уровень серотонина в тромбоцитах у больных 1 группы является достаточно показательным результатом. Содержание внутриклеточного серотонина отражает устойчивые тенденции уровня захвата на протяжении достаточно длительного промежутка времени. Таким образом, факт снижения концентрации нейромедиатора указывает на низкий уровень интенсивности его захвата пресинаптическими структурами, что в принципе является распространенным феноменом для контингента больных с психическими расстройствами. Проявления гипофункции транспортных индоламиновых механизмов в виде снижения плотности имипраминовых рецепторов тромбоцитов, уменьшения максимальной скорости захвата и снижения концентрации серотонина в тромбоцитах в той или иной форме обнаруживаются при депрессивных, обсессивно-компульсивных, панических расстройствах (Dubovsky S.L., 1994; van Praag H.M., 1994). Результаты наших исследований показали, что низкий уровень внутриклеточного серотонина характерен для ряда депрессивных синдромов и психопатий тормозимого круга,(Дроздов А.З.,1996), и в этом смысле больные 1 группы являются примером общей неспецифической тенденции изменений нейрохимического статуса индоламиновых систем при психических расстройствах. Таблица 7.5. Состояние индоламиновой системы при парафилиях у испытуемых с нарушениями половой идентичности.
* — p * — и ** — p (данные по группе 4 не представлены вследствие ее малочисленности) Несмотря на то, что у пациентов 1 группы содержание серотонина во внеклеточной жидкости (плазме) не отличается от контрольного, средние значения данного показателя у представителей других групп превышают контрольные значения, что особенно ярко проявляется во 2 группе (табл 7.5). Подобная направленность изменений содержания серотонина в плазме крови также показывает, что процессы захвата могут быть снижены у лиц с нарушениями половой идентичности. Таким образом, результаты недвусмысленно указывают на тенденцию к понижению скорости захвата серотонина при парафилиях. По нашему мнению, данный процесс является приспособительным изменением физиологии медиаторных систем в условиях недостаточности моноаминовых постсинаптических функций (Дроздов А.З.,1996). Еще одна общая тенденция, характерная для всех групп, заключается в значительном разбросе исследуемых показателей. Эта тенденция проявляется и в значениях уровня серотонина плазмы, серотонина тромбоцитов, Vмах. Иллюстрацией данного положения является рис.7.2. Само по себе подобное проявление дисбаланса периферических индоламиновых систем может рассматриваться как доказательство нарушений нормального функционального состояния центральных серотониновых структур. Существенное увеличение разбросов изучаемых показателей по мере нарастания клинических признаков нарушения половой идентичности свидетельствует об определенной их взаимосвязи с неустойчивым состоянием центральных моноаминовых механизмов. Обращает на себя внимание (рис.7.2) тот факт, что при увеличении степени выраженности нарушений половой идентичности обнаруживается очевидный рост числа пациентов, у которых фиксировались высокие значения содержания серотонина в тромбоцитах. Можно предположить, что увеличение поглощения нейромедиатора из зоны взаимодействия с рецепторами является одним из патогенетических механизмов, формирующих изменение концентрации серотонина в синаптической щели, а это, в свою очередь, может влиять на модифицикацию половой идентичности человека.
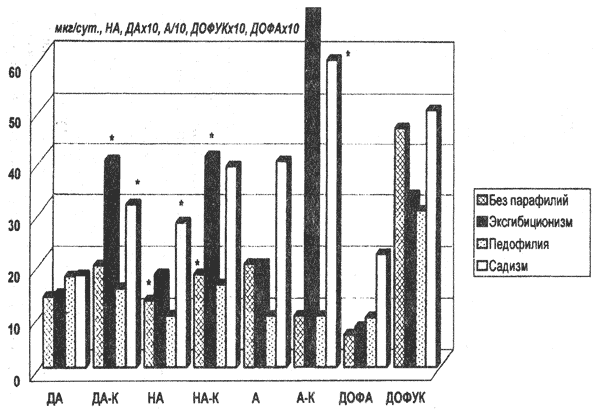
В настоящее время хорошо известно, что соотнесение патологического функционального состояния индоламиновых систем с какой-либо конкретной психической патологией не даёт каких-либо практических результатов (Poldinger W. T et al. 1991; van Praag H.M., 1994). С другой стороны, широкий ряд психопатологических состояний сопровождается нарушениями серотониновой медиации (Dubovsky S.L., Thomas M., 1995; van Praag H.M., 1994). Более того, препараты серотонинового типа действия являются основными терапевтическими средствами при депрессивных, тревожно-фобических, обсессивно-компульсивных, панических расстройствах (Aberg-Wistedt A.,1989; Dubovsky S.L., Thomas M., 1995). Современные теоретические представления достаточно адекватно интерпретируют имеющиеся клинические данные. Очевидно, что универсальные тормозные системы головного мозга, одними из представителей которых являются индоламиновые, обеспечивают общую регуляцию функционального статуса нейрональных систем мозга, нарушения которой проявляются в изменении множества видов конкретных поведенческих реакций. Не является исключением и половое поведение, различные аспекты которого в достаточной степени меняются при воздействиях на серотонинергические системы. Данные настоящего исследования показывают, что клинические признаки нарушений половой идентичности не обнаруживают прямой связи с четко направленными перестройками серотониновых механизмов. Вместе с тем по мере усиления выраженности данного расстройства в популяции больных отчетливо проявляются признаки дисбаланса серотонинового метаболизма, очевидно отражающего наличие разнонаправленных тенденций в уровне функционирования аппарата индоламиновой медиации. Таким образом, попытка соотнесения клинических признаков нарушений половой идентичности, механизмы реализации которых находятся на высоком уровне организации высшей нервной деятельности, с патологией неспецифической модулирующей системы головного мозга привела к результатам, однозначная интерпретация которых в настоящее время весьма затруднительна. По всей вероятности патология, серотониновых структур (также как и катехоламиновых) представляет основу развития разнообразных дизонтогенетических состояний, затрагивающих различные этапы психосексуального развития (половое самосознание и психосексуальные ориентации), поскольку моноамины в значительной мере определяют половую дифференцировку головного мозга в эмбриогенезе (Резников А.Г.,1982), участвуют в становлении психосексуальности и модификации полового поведения. С этой точки зрения результаты настоящей работы, свидетельствующие о явном дисбалансе серотониновой нейромедиации у лиц с нарушениями половой идентичности, трудно переоценить. В связи с этим необходимо продолжать целенаправленный поиск структурных клинических закономерностей в рамках аномального сексуального поведения, появление и развитие которого имеет более отчетливую связь с проявлениями нарушений функциональной активности моноаминовых медиаторных систем. 7.3.3 Особенности психопатологической картины и нейромедиаторыКак известно, нарушения обмена моноаминов обнаружены при различных психических расстройствах, в том числе психопатиях, депрессивных состояниях, алкоголизме и наркоманиях (Дмитриева Т.Б. 1990; Коган Б.М., Анохина И.П., 1990).Не вызывает сомнений факт, что патология моноаминового обмена является одной из причин развития указанных состояний (van Praag H.M. et al., 1990). Тем более это справедливо для парафилий, т.е. тех состояний измененного сексуального влечения, в основе которых лежат специфические нейрофизиологические, нейрохимические и иные биологические детерминанты, связанные с патоорганическими, нейрогуморальными и психологическими особенностями таких лиц. Поскольку моноаминовые медиаторные системы контролируют гипоталамические и гипофизарные механизмы, регулирующие синтез половых гормонов, половую дифференцировку и развитие первичных и вторичных половых признаков, генетическая измененность (прежде всего из-за нарушений ферментативной деятельности) тех или иных стадий метаболизма катехоламинов может быть связана с проявлениями аномальных сексуальных реакций. Каковы бы ни были теории аномального сексуального поведения — от культуральных до биомедицинских — все они в конечном итоге вынуждены учитывать внутреннюю структуру переживаний и форму, в которой эти переживания проявляются, существенно различающуюся даже при внешне однотипных девиантных действиях. Так, одна из классификационных систем, основанная на мотивационном спектре стимулов — от исключительно экзогенных до исключительно эндогенных, подразумевает существование 1) социогенного, 2) ситуационного, 3) импульсивного, 4) кататимного и 5) компульсивного сексуального насилия. Использование в данном контексте дефиниции «компульсии» соответствует представлениям, отраженным в DSM-III-R, о том уровне непроизвольности перверсных эго-дистонических переживаний, когда правомерно говорить о патологии. Непосредственно связанные с обсессиями компульсии представляют собой ритуальные поведенческие акты, направленные на редукцию и нейтрализацию тревоги, вызываемой навязчивыми переживаниями. Исключающими диагноз обсессивно-компульсивного расстройства являются озабоченность мыслями о еде у пациентов с расстройствами аппетита, компульсивные действия, приводящие к получению удовольствия (такие, как использование наркотиков, игра в азартные игры и другие виды аддиктивного поведения), а также сексуальные поведенческие акты. Однако все пять критериев диагностики обсессивно-компульсивного расстройства оказываются пригодными и для квалификации тех насильственных и непреодолимых сексуальных действий, которые также обозначаются как компульсивные. Эти переживания и действия 1) могут занимать длительное время (больше часа в день), 2) вмешиваются и препятствуют функционированию, 3) вызывают выраженный дистресс, 4) остаются упорными и 5) трудно поддающимися контролю индивида. Поэтому отделение их от собственно обсессивно-компульсивных расстройств, которое обычно принято, выглядит несколько условным с клинической точки зрения. Для диагностики парафильного поведения вполне пригодны и критерии, предлагаемые для квалификации «расстройств контроля импульса». Феноменологические исследования лиц с различными вариантами «расстройств контроля импульса» показали тесную их связь прежде всего с тревожными, в особенности с обсессивно-компульсивными расстройствами. Более того, сами по себе патологические импульсы и следующие за ними действия имеют сходство с обсессиями и компульсиями, в связи с чем, например, клептомания, пиромания и трихотилломания могут рассматриваться как формы обсессивно-компульсивного расстройства. Специальных работ, посвященных биохимическому исследованию проявлений «расстройств контроля импульса» в настоящее время немного. Имеются сведения о том, что у лиц с импульсивным агрессивным или аутоагрессивным поведением и с пироманиями снижено содержание метоксифенилэтиленгликоля (МОФЭГ) в ликворе по сравнению со здоровыми людьми. У лиц с патологической страстью к игре в карты увеличена экскреция норадреналина и содержание МОФЭГ в ликворе и снижена концентрация МОФЭГ в крови по отношению к контрольным показателям. Особый интерес при изучении биохимических основ патогенеза обсессивно-компульсивных расстройств вызывает состояние серотониновой медиаторной системы, поскольку ингибиторы обратного захвата серотонина являются эффективным средством купирования навязчивостей (Karakiulakis G. et al., 1978.). Однако работы, посвященные биохимическому исследованию проявлений «расстройств контроля импульса», пока немногочисленны и противоречивы. Показано, что у лиц с импульсивным агрессивным или аутоагрессивным поведением и с пироманиями снижены содержание 5-оксииндолуксусной кислоты (основного метаболита серотонина в организме, 5-ОИУК) в ликворе и скорость захвата меченого серотонина тромбоцитами [12] по сравнению со здоровыми людьми. В ряде работ авторы не нашли различий в скорости захвата серотонина тромбоцитами у здоровых лиц и у больных с обсессивно-компульсивными расстройствами (Karakiulakis G. et al., 1978.). У этих же больных обнаружено увеличение содержания 5-ОУИК в ликворе, что может отражать увеличение кругооборота серотонина в мозге с увеличением обратного захвата серотонина пресинаптическими окончаниями. Следовательно, эти данные соответствуют факту клинической эффективности блокаторов обратного захвата серотонина при обсессиях (Karakiulakis G. et al., 1978.). Таким образом, результаты проведенных исследований, доказывающих неразрывность описанных расстройств, с непреодолимыми импульсами или побуждениями к совершению специфических бессмысленных или вредных поступков, приводят одновременно к утверждениям, что в основе такой общности могут лежать биологические нарушения, единые для спектра аффективных расстройств, что диктует необходимость изучения состояния моноаминовых систем при этих патологических проявлениях. Поскольку средние цифры показателей обмена катехоламинов в общей группе лиц с парафилиями достоверно не отличаются от контрольных значений, мы проанализировали распределение исследуемых параметров моноаминового метаболизма у испытуемых с парафилиями, объединенных по различным клиническим признакам. Обнаружить зависимость биохимических показателей от нозологической квалификации больных с парафилиями, от объекта сексуальной направленности пациента не удалось. Наиболее выраженными были биохимичесие различия у испытуемых без компульсивных расстройств 1 группа и у лиц с компульсивными формами парафилий 2 группа. В ходе клинического исследования среди лиц, совершивших противоправные сексуальные деяния, была выделена группа испытуемых, у которых парафильные акты носили выраженные черты компульсивности. У одного из обследованных девиантный акт имел признаки пароксизма. По другим характеристикам вошедшие в группу испытуемые не отличались от остальных испытуемых с расстройствами сексуального влечения. Не было отмечено какой-либо специфики в квалификации иных психических расстройств, среди которых преобладали различные варианты органического поражения головного мозга. В результате проведенного биохимического исследования крови и мочи испытуемых обнаружены существенные отличия в показателях, характеризующих состояние катехоламиновой системы, у представителей сравниваемых групп. Так, содержание свободных и, особенно, конъюгированных форм НА, ДА и ДОФУК достоверно выше у представителей 2 группы. Особенно значимы эти различия при рассмотрении суточной экскреции изучаемых веществ (табл.7.6, рис.7.3). У этих же больных выявляются низкие значения коэффициента ДА/ДОФУК (табл.7.7), который в определенной степени отражает состояние процесса окислительного дезаминирования ДА, в основном происходящее в пресинаптических терминалях под влиянием МАО после обратного захвата нейромедиатора из синаптической щели. Низкие значения соотношения ДА/ДОФУК в суточной моче у испытуемых 2 группы свидетельствуют о высокой интенсивности данного катаболического пути. Подобная биохимическая особенность является следствием либо высокой активности МАО (ферментной системы окислительного дезаминирования моноаминов в организме), либо большой концентрации медиаторных молекул в пресинаптических терминалях, что, в свою очередь, может быть обусловлено высокой скоростью обратного захвата медиаторов из синаптической щели. Действительно, согласно полученным данным, интенсивность захвата меченого серотонина тромбоцитами у лиц с проявлениями компульсивности при совершении девиантных действий достоверно выше, чем в противоположной группе (табл.7.9). Как известно, тромбоциты рассматриваются в качестве модельного объекта центральных пресинаптических окончаний, что дает основание предполагать наличие аналогичных процессов в ЦНС. Таблица 7.6. Экскреция свободных и конъюгированных форм катехоламинов, свободных форм ДОФА и ДОФУК при парафилиях
* — р ** р *** p
Таблица 7.7. Значения коэффициента ДА/ДОФУК в моче при компульсивных расстройствах у больных с парафилиями.
* — р Другой путь физиологической инактивации биологически активных моноаминов у человека — их конъюгация с остатками неорганических кислот, происходящая в печени, тромбоцитах и ЦНС, главным образом по пути образования сульфо-О-эфиров. Эта сторона метаболизма моноаминов в приложении к психическим расстройствам практически не изучалась, хотя физиологическая роль сульфоконъюгирования достаточно велика. Как уже отмечалось, выделенные группы разительно отличаются по содержанию свободных и связанных форм катехоламинов (рис7.3) и ДОФУК. У испытуемых 2 группы количество связанных форм намного выше, чем в первой группе, также как и интенсивность процессов конъюгирования, оцениваемая по соотношению свободных и конъюгированных форм (табл.7.8). Если вспомнить далеко не бесспорное предположение о том, что уровень сульфоконъюгатов ДОФУК в моче может отражать степень ДА-нейрональной активности в ЦНС, то не исключено усиление функциональной активности этой нейромедиаторной системы в соответствующих областях мозга испытуемых. Таблица 7.8. Значения коэффициента свободные/конъюгированные формы для КА и ДОФУК в моче при компульсивных расстройствах у больных с парафилиями.
Аналогичные результаты получены при обследовании лиц с иными компульсивными расстройствами (пиромания, страсть к азартным играм и другие). Авторы показали, что у обследованных лиц экскреция НА выше, по сравнению с контрольной группой. Вообще, в немногочисленных работах, посвященных оценке уровня функционирования моноаминовых систем при «расстройствах контроля импульса», авторы находили ту или иную патологическую нейрохимическую составляющую. У двух обследованных групп имелись также определенные различия между параметрами, характеризующие состояние периферической индоламиновой системы (табл.7.9). Так, у лиц с компульсивной формой парафилий увеличена скорость захвата серотонина тромбоцитами по сравнению с другой группой. При этом значения Км процесса захвата в этих группах не различались. Содержание серотонина в тромбоцитах также не различалось у двух групп, а уровень серотонина в плазме крови имел устойчивую тенденцию к снижению в группе испытуемых с компульсиями, что соответствует данным по захвату тромбоцитами серотонина. Принимая во внимание модельный характер процессов в тромбоцитарных клетках для центральных серотониновых механизмов, можно допустить, что у лиц с компульсивными девиантными актами увеличена скорость обратного захвата серотонина пресинаптическими окончаниями. Основываясь на косвенных данных, мы полагаем, что это увеличение свидетельствует об относительной недостаточности серотонинергических процессов. Так, имеются данные об эффективности ингибиторов обратного захвата серотонина при обсессивно-компульсивных расстройствах (Karakiulakis G. et al., 1978.), что косвенно подтверждает наше предположение о слабости индоламиновых механизмов. Известно, что основные индоламиновые системы мозга являются тормозными регулирующими системами. Очевидно, что недостаточность тормозного контроля некоторых нейронных образований может иметь отношение к формированию клинических проявлений обсессивно-компульсивных расстройств. Таблица 7.9. Параметры захвата 3Н-серотонина тромбоцитами при компульсивной и некомпульсивной формах парафилий
* — р Основываясь на полученных данных, можно предположить, что у лиц с парафильными компульсивными расстройствами более выражена функциональная недостаточность основных тормозных систем (норадреналиновой и серотониновой) головного мозга по сравнению с лицами, совершавшими девиантные акты и у которых не было выявлено компульсивных расстройств. Тезис о недостаточности тормозных структур мозга подтверждается работами, в которых авторы показали увеличение углеводного метаболизма в различных отделах мозга у больных с обсессивно-компульсивными расстройствами. Слабость процессов торможения при сложившемся очаге насильственных побуждений может приводить к невозможности заблокировать развертку тотальной спонтанной активности нейронов, что в числе прочего ведет к гипервозбуждению и невозможности контролировать свои действия в момент совершения эксцессов, отвечающих требованиям доминантного нейрофизиологического образования, особенно сильно выраженным в случае сексуального влечения. «Расстройства контроля импульса» характерны для ряда нервно-психических расстройств, имеющих общие клинико-биохимические проявления, поэтому не исключено, что обнаруженные в настоящем исследовании нейрохимические закономерности связаны в первую очередь не с наличием парафилий, а с феноменом компульсивности. Очевидно, что особое состояние сознания в определенные моменты не является типичным расстройством, характерным только для парафилий. В данном случае уникальным является сам характер насильственного импульса, формируемого на основе сочетания биологических свойств нейронных систем головного мозга (врожденных или приобретенных) и специфических онтогенетических предпосылок. При анализе полученных данных возникает вопрос, в какой степени особенности индоламиновых механизмов влияют на развитие сексопатологических особенностей испытуемых, или, в более общем плане, в какой степени связан феномен компульсий с формированием парафилий? Полученные клинические данные показывают, что у всех лиц с компульсивным поведением (суть которого составляют такие феномены, как стереотипизация, «клиширование», потеря произвольности и волевого контроля над побуждениями) в основе патологических побуждений лежат аутохтонные механизмы формирования, связанные с конституционально-дизонтогенетической базой нарушения психосексуальной ориентации. Таким образом, очевидно, что между «расстройствами контроля импульса» и парафилиями существует определенная внутренняя связь. Упомянутая выше внутренняя взаимосвязь между «расстройствами контроля импульса» и парафилиями может получить объяснение с учетом важнейшей роли моноаминовых систем в формировании половой дифференциации мозга в пренатальном периоде. Будучи универсальными регуляторами функций взрослого организма катехоламины и индоламины являются так же эмбриональными гормонами, действие которых начинается с ранних этапов дробления оплодотворенной яйцеклетки. На более поздних стадиях развития моноамины в значительной мере определяют половую дифференцировку головного мозга. В качестве одного из ключевых звеньев регуляторного воздействия по переключению индивидуального развития с фемининного на маскулинный выступает достаточная активность моноаминовых систем в определенных отделах развивающегося мозга. Согласно, основную роль в моноаминергическом регулировании формирования половых признаков играет норадреналин и значительно меньшее участие принимают серотонин и дофамин. Исходя из этих данных, можно предположить, что различия в метаболизме катехоламинов между подгруппами испытуемых с парафилиями могут быть обусловлены разным уровнем функционирования норадренергических систем головного мозга, ядра которых локализованы в среднем и продолговатом мозге [17], а терминали оканчиваются практически во всех вышестоящих отделах мозга, в том числе и гипоталамусе, который является центром регуляции нервно-гуморальных взаимоотношений. Клиническая картина дает основание полагать, что у лиц с признаками парафилий оказывается неполноценным процесс становления маскулинных нейрогуморальных механизмов. Полученные результаты свидетельствуют о том, что у лиц с парафилиями усилен периферический метаболизм катехоламинов по сравнению с группой лиц, совершивших девиантные сексуальные акты, но в меньшей степени проявляющих аномальное сексуальное влечение. Можно предположить, что этот факт отражает относительную слабость центральных адренергических механизмов у этой первой группы лиц. Нельзя исключить, что недостаточность катехоламиновых механизмов проявлялась и на ранних стадиях онтогенеза, что привело к определенным деформациям регулирующих сигналов, определяющих нормальное формирование комплекса психосексуальных реакций. Таким образом, параллельное развитие дизонтогенетических расстройств и обсессивно-компульсивных нарушений у лиц с парафилиями в значительной степени может быть обусловлено комплексом нарушений моноаминовых механизмов в раннем эмбриогенезе. 7.3.4. Состояние нейромедиаторных систем при различных вариантах парафилийНормальный постнатальный онтогенез, включающий в себя и нормальное сомато- и психосексуальное развитие, возможен в том случае, если нормально протекают процессы эмбрионального внутриутробного развития с правильным становлением мозговых структур, ответственных за аффективно-когнитивные процессы. Серотонинергическая иннервация актуализирует внутрикорковую интеграцию, обеспечивающую возвратное и/или латеральное торможение. Этот вид торможения регулирует эффективную функцию нейронов коры и формируется в онтогенезе одним из последних. Аминергические системы мозга осуществляют консолидирующее влияние на нейроны неокортекса. Нарушение ядер шва в 30-дневном возрасте у котят вызывает бессонницу, появление высокой двигательной активности (аналог гипердинамического синдрома), значительную и постоянную активность электрокортикограммы, что указывает на нарушение тормозных механизмов. В результате разрушения мезокортикальных дофаминергических нейронов у крыс обнаружено снижение уровня исследовательской деятельности. Инъекция апоморфина (агонист дофамина) способствовала восстановлению исследовательского поведения (Fink J.,Smith G., 1980). При инъекции крысам 6-OHDA в мезолимбическую дофаминергическую систему приводило к ухудшению обучения. Можно предполагать, что баланс между этими двумя важными системами, в реализации аномального сексуального агрессивного поведения легко нарушается под влиянием различных патогенных факторов, например, действующих во внутриутробном периоде, в интранатальном раннем постнатальном и в дальнейшем онтогенезе индивидуума. Необходимо обратить внимание на то, что медиаторные системы в онтогенезе формируются разновременно, их развитие протекает неградуально, т.е.могут быть случаи замедления их развития, рано созревшие системы выполняют множественные функции, связанные не только с синаптической передачей, но и с регуляцией других процессов онтогенетического развития; что отчасти оказывается возможным благодаря избыточному созреванию данных систем (Раевский В.В,1991). В регуляции агрессивного поведения участвуют практически все известные нейромедиаторы, однако наибольшее внимание следует уделить катехоламинэргической системе (дофамин) и индоламиновой (серотонин). Так, в опытах на крысах было отмечено, что торможение синтеза катехоламинов А-метилпаратирозином (АМПТ) снижает выраженность агрессивной реакции. Таким же образом действовал и ингибитор тирозингидроксилазы. Подавление рецепторов дофамина галоперидолом также вызывало снижение агрессивности у лабораторных животных. Использование агонистов и прекурсоров КА — систем также показало важную роль этих систем в интеграции агрессивного поведения. Высокий уровень агрессивности наблюдался у мышей, которые содержались на диете с добавлением прекурсора дофамина L-ДОФА. Фенилалкиламины (фенамин, метамфетамин (первитин), усиливающие дофаминергическую передачу, вызывали нарушения в эмоционально-мотивационной сфере — агрессивность, страх, гиперсексуальность (Отеллин В.А., Арушанян Э.Б.,1989). Введение ингибитора серотонина (парахлорфенилаланина) самцам крыс вызывало половую активацию не только у относительно инертных животных, но и у крыс с повышенной половой активностью. Причем усиливались не только признаки гетеросексуального поведения, но и половая мотивация: крысы становились способными совершать садки не только на самок, находящихся в состоянии течки, но и на нерецептивных самок (Sjoerdsma et al., 1970). Результаты опытов, полученных многими исследователями, которые применяли для анализа ингибитор синтеза серотонина — парахлорфенилаланин привели к однозначному выводу об ингибирующей роли серотониновых механизмов мозга на мужское половое поведение. Ряд авторов полагают, что влияние серотонина головного мозга на половое поведение самцов тесно связано с состоянием семенников. При опытах с использованием двух групп животных — половозрелых интактных и кастрированных самцов — которым вводили парахлорфенилаланин, авторы обнаружили, что у интактных самцов признаки полового возбуждения наблюдались почти в 30% случаев, в то время как ни у одного представителя другой группы ни в одном случае признаков полового возбуждения не отмечалось. Введение интактным животным одного тестостерона вызывало активацию полового поведения у меньшего числа животных, чем введение парахлорфенилаланина. В то время как у крыс, получавших тестостерон в комбинации с парахлорфенилаланином, появлялись признаки гиперсексуальности, даже у кастрированных самцов половая активность отмечалась в значительно большем проценте случаев, чем у интактных крыс, получавших один тестостерон. Внутрибрюшинное введение интактным или кастрированным самцам, получавшим парахлорфенилаланин и тестостерон, 15 мг 5-окситриптофана полностью блокировало на 2-3 ч. половую активность. Из этого можно сделать вывод, что половое поведение, активируемое у самцов крыс тестостероном, угнетается серотонином на уровне головного мозга; для проявления полового поведения недостаточно одного падения уровня серотонина (или тонуса серотонинергических нейронов). Для этого обязательно наличие тестостерона. Эти данные получили свое подтверждение в работах других исследователей. Имеются предположения, основанные на экспериментах, позволяющие делать выводы о том, что в головном мозгу серотонин ингибирует структуры, чувствительные к тестостерону и связанные с активацией мужского типа поведения. Нельзя также исключить той возможности, что у самцов тестостерон оказывает угнетающие воздействие на серотонинергические механизмы. Сексуальную активность кастрированных самцов крыс поддерживали на субмаксимальном уровне небольшими дозами тестостерона, введение таким животным парахлорфенилаланина резко усиливало половое поведение; процент садок повысился с 49 до 90% (Malmnas C.O., Meyerson B.J., 1971; Malmnas C.O.,1974). На сегодняшний день можно считать установленным, что в агрессивности самцов большую роль играет фукциональная активность мужских половых желез (Fredericson E., 1950; Levy J.V., King J.A., 1953; Edwards D.A., 1969; Никулина Э.М., 1976), которая в агрессивности хищника особой роли не играет. Сексуальную агрессию можно считать одной из форм внутривидовой агрессивности. Имеются немногочисленные данные об участии серотонина в механизмах реализации внутривидовой агрессии. Прежде всего показано различное содержание серотонина в стволе мозга у мышей различных линий, отличающихся по агрессивности согласно теста внутривидовой агрессии самцов (Maas, 1962), причем более высокое содержание у неагрессивных животных отмечено и в опытах с направленной селекцией на агрессивность и на неагрессивность (Langerspetz K.M., Langerspetz K.Y.,1974). Блокада синтеза серотонина парахлорфенилаланином значительно усиливала внутривидовую агрессивность у котов (Ferguson J. et al.,1970; Mc Donnell M.F. et al.,1971). К заключению о зависимости агрессивности самцов от баланса между серотонином мозга и дофамином привели опыты на мышах, инфицированных вирусом Herpes vulgaris (Lycke E. et. al.,1969). Данный вирус вызывал у мышей энцефалит, приводящий к понижению синтеза серотонина и повышению синтеза дофамина. Инфицированные мыши после введения им H69/17 проявляли отчетливо повышенную агрессию, чего не происходило у обычных мышей. Можно предполагать, что в реализации аномального агрессивного поведения играет роль не столько абсолютное снижение серотонина как ингибирующего данное поведение фактора, сколько баланс между катехол- и индоламиновыми системами. Импульсная активность нейронов миндалины резко уменьшается после парентерального введения 5 окситриптофана. Имеются данные, указывающие на различное содержание серотонина в мозге мышей различных линий, которые отличают по степени внутривидовой агрессии самцов. Причем более высокое содержание серотонина отмечалось у неагрессивных самцов. Блокада синтеза серотонина парахлорфенилаланином заметно усиливала внутривидовую агрессию у котов, а введение в вентромедиальный гипоталамус серотонина снижало агрессивность у кошек. Снижение на 50% уровня серотонина, происходившее после разрушения ядер шва, приводило к тому, что агрессивность самцов увеличивалась почти вдвое. В 1976г. Asberg et al. было обнаружено, что у депрессивных больных со сниженной экскрецией ГИУК (основного метаболита серотонина) достоверно было увеличено как число суицидов (аутоагрессия), так и количество актов агрессии, направленной вовне. Итак, серотонин представляет собой фактор (нейромедитор), достоверно снижающий агрессивность (Попова Н.К., Науменко Е.В., Колпаков В.Г., 1978). Ранняя функция серотонинергических нейронов не связана с синаптической передачей. Предполагается, что они регулируют пролиферацию нейронных предшественников и их миграцию. Возможно, серотонинергические волокна, подходящие к клеткам-мишеням, играют по отношению к ним роль дифференцировочного сигнала. Дифференцировка гранулярных клеток в некоторых отделах мозга (мозжечок и гиппокамп) начинается после появления в этих зонах серотонинергических терминалей (Baker P.C., Quay W.B.,1969; Buznikov G.A. et al., 1970; Wallace J.A.,1982). Любое патогенное воздействие в раннем онтогенезе может легко нарушить нормальную функцию серотонинергической системы и тем самым вызвать нарушение созревания других нейромедиаторных систем и структур мозга, что в конечном счете приведет к их дисфункции и к ухудшению общей интрацентральной интеграции, без которой нормальное функционирование головного мозга невозможно (.Раевский В.В,1991). Исследование нейрохимических основ патологического поведения способствует раскрытию не только содержательной его стороны, но в значительной степени — и клинического его своеобразия, например, того стереотипного характера, который является одним из фундаментальных механизмов становления парафилий. В последние годы экспериментальное подтверждение получило положение о жесткой зависимости реализации врожденого поведения или выработанных при жизни способов действий от уровня активности медиаторных систем. Так, J.Fink, G. Smith (1980) показали, что для того, чтобы врожденное или условно-рефлекторное поведение осуществилось, требуется некоторый минимальный уровень активности медиаторных систем мозга, при снижении активности дофаминергической системы ниже критического уровня животное вообще не способно к какому-либо мотивированому поведению. Были предприняты попытки изучения механизмов действия и влияния медиаторных систем и веществ, в них участвуюших, на поведение в целом. Например, фенамин, введенный животным в больших дозах, вызывает монотонные реакции (принюхивание, насильственный грумминг и т.д.) и резко снижает активность на внешние сигналы. У человека при приеме фенамина в дозе 10 мг/кг тоже могут возникать стереотипии мелкие движения головы, конечностей (Ильюченок Р.Ю.,1972). В результате опытов были сделаны следующие выводы: активирование дофаминергических структур, так же как и ингибирование, приводит к расстройству функции обобщения и абстрагирования у интактных кошек. Избыток, равно как и недостаток, нейромедиатора дофамина в ЦНС одинаково неблагоприятен для протекания высших психических процессов. Переход за физиологические пределы дофаминергической передачи неминуемо грозит поражением функции обобщения. В дальнейшем, после исследования холинергической системы было выяснено, что ингибирование холинреактивных структур центральными холинолитиками (атропином или скополамином) не сказалось существенным образом ни на протекании условно-рефлекторной деятельности, ни на решение задач, на обобщение, отвлечение разной степени сложности (Мухин Е.И., 1990). После многочисленных опытов с медиаторными системами ученые пришли к выводу о том, что непосредственное участие в обеспечении интеллектуальных операций принимает в основном дофаминергическая система. Также были предприняты попытки изучения как участия нейромедиаторных систем в образовании простых условных рефлексов, так их участие в более сложной интеллектуальной деятельности. Можно говорить о том, что, с одной стороны, есть механизм образования стереотипий или стереотипного поведения, который основан на механизме образования условных рефлексов, с другой стороны есть механизм образования новых ассоциативных связей. Эти два механизма находятся в антагонизме по отношению друг к другу. Механизм образования новых ассоциаций приводит к выделению новых значимых признаков и лежит в основе абстрагирования, «абстрактного мышления» и творчества, в то время как механизм образования стереотипий стремится к упорядочиванию уже известных признаков и связей между ними. Учитывая полученные данные по участию медиаторных систем в обучении, можно предположить, что в механизме создания стереотипий участвуют все три медиаторные системы, а в механизме создания «нового» будет участвовать преимущественно дофаминергическая система и соответствено, при ее нарушении будет больше возможностей образования стереотипий и стереотипного поведения. Возможно, что нарушение взаимодействия систем, их дисбаланс, возникающий при поломках какой-либо одной системы и выражаюшеся в дезадаптации, это и есть проявление несовершенства нейрохимических механизмов. В целом полученные данные о патобиологических основах расстройств сексуального поведения позволяют подойти к формулировке целостных патогенетических концепций парафилий, в том числе их агрессивных вариантов. Так, Flor-Henry (1989) считает, что дофамин- и серотонинергическая системы имеют билатеральные проекции к лимбико-кортикальным структурам (гиппокамп, миндалина, септум и фронтальный неокортекс) и именно эта билатеральность обусловливает воздействие межполушарной асимметрии на регуляцию сексуального поведения. Нейротрансмиттерные системы находятся в антагонистических взаимоотношениях и организованы билатерально асимметрично. Наиболее очевидные результаты биохимического исследования крови у испытуемых с парафилиями обнаруживаются у испытуемых с различной степенью агрессивности. Условность трактовки агрессивности не помешала выявить ряд интересных закономерностей. В частности, уровень ДОФА и конъюгированных форм ДА в крови у лиц без парафилий был значительно ниже, чем у больных с парафилиями практически независимо от типа парафилии и направленности агрессивных проявлений (табл.7.10). Таблица 7.10. Содержание свободных и конъюгированных форм КА, а также ДОФА и ДОФУК в крови испытуемых с парафилиями
* — р + — р $$ — р Поскольку содержание НА и моноаминовых молекул в последующих стадиях метаболизма КА не отличалось в описываемых группах, с достаточной степенью уверенности можно утверждать, что у лиц с нарушениями полового влечения по типу эксгибиционизма, педофилии, садизма синтез катехоламинов значительно усилен по сравнению с лицами без признаков наличия парафилий. Весьма примечательно, что у данных групп отмечаются сходные паттерны экскреции (с описанными для крови) катехоламиновых молекул (табл.7.11). Так, экскреция свободных форм НА, конъюгированных молекул НА, А, ДА у больных с парафилиями в ряде случаев значительно превышает показатели у испытуемых без парафилий (рис.7.4). Итак, принципиальная схожесть периферических показателей крови и мочи в моделях парафильного агрессивного поведения позволяет сделать вывод о значительном ускорении метаболизма катехоламиновых медиаторов у лиц с проявлениями истинных парафильных тенденций.
По нашему мнению, обнаруженная тенденция усиления метаболизма КА с увеличением в первую очередь синтеза катехоламинов у больных с парафилиями на фоне кругооборота КА у лиц без признаков нарушения полового влечения, имеет неспецифический характер по отношению к этиологии и патогенезу различных форм парафилий. В некоторых случаях обнаруженные особенности могут отражать наследуемые качества катехоламиновых структур, необходимые для фенотипического проявления поведенческих ассоциатов парафильных расстройств. Для обоснованного утверждения о более интимном участии моноаминовых систем в формировании и развитии нарушений полового влечения необходимы дальнейшие клинико-биохимические исследования. Таблица 7.11. Экскреция свободных и конъюгированных форм катехоламинов, свободных форм ДОФА и ДОФУК при парафилиях
* — по сравнению с группой 3 (один символ — р+ — по сравнению с группой 0 Таким образом, мы обнаружили факт существования взаимосвязи между общей активностью катехоламиновых систем на периферии и клиническими проявлениями парафилий. Не исключено, что полученные данные свидетельствуют в пользу двуединой модели парафильного поведения, в рамках которой условия, определяющие его формирование, могут включать как специфические для парафилий этиопатогенетические звенья, приводящие к аномальному поведению в отсутствие иных психических расстройств, так и неспецифические, в том числе и психопатологические факторы. На наш взгляд, именно соотношение «специфического» и «неспецифического» во многом определяется индивидуальными особенностями организации катехоламиновых систем, и указывает на патогенетическое значение нарушений нейрохимических процессов в формировании и развитии аномального сексуального поведения. Проведенный комплекс нейрохимических исследований показал, что у больных парафилиями обнаруживается целый комплекс нарушений катехоламинергической и индоламинергической нейромедиации, особое место в котором занимают нарушения процессов сульфоконъюгирования катехоламинов и обратного захвата моноаминов. В то же время полученные результаты выявляют чрезвычайную гетерогенность организации нейромедиаторных процессов у данного контингента больных и ставят под сомнение принципиальную возможность проведения индивидуальной патогенетически обоснованной фармакотерапии больных, ориентируясь лишь на диагноз: Парафилия. Несомненно единственно правильными будеу подходы к попытке купирования отдельных психопатологических синдромов в рамках парафилий и создание новых схем фармакологического воздействия на конкретные звенья метаболизма моноаминовых нейромедиаторов у каждого пациента, исходя из результатов исследования состояния основных механизмов, определяющих функциональную активность нейромедиаторных моноаминовых систем организма. Литература
Аномальное сексуальное поведение(Под редакцией А. А. Ткаченко. М., 1997)
|